Transcription occurs in three steps: (i) the initiation step, in which RNA polymerase II (Pol II) binds to the promoter region of a gene and starts RNA synthesis, (ii) the elongation step, in which Pol II actively synthesizes RNA in a 5’-to-3’ direction, and (iii) the termination step, in which Pol II reaches the termination site and is released from DNA. These steps are accomplished by numerous factors that bind to DNA or Pol II. Turning the clock back to the 1990s, research on the transcription initiation stage was in full bloom, and it was widely believed that the transcription initiation stage was the key to on/off regulation of gene expression in eukaryotes.
Under these circumstances, our laboratory turned its attention to DRB, a specific inhibitor of RNA polymerase II transcription, which, when added to cultured mammalian cells, was reported to inhibit the synthesis of long transcripts and cause the accumulation of short transcripts. Thus, it was hypothesized that DRB might inhibit the transcription elongation step. Surprisingly, however, DRB did not inhibit purified RNA polymerase II. We therefore sought to elucidate the mechanism of DRB-mediated transcription inhibition using a biochemical approach.
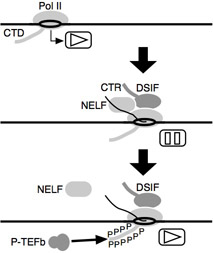
As a result, we identified two new transcription elongation factors, DSIF (SPT4-SPT5) and NELF. We also found that P-TEFb, a transcription elongation factor discovered by David Price’s lab, works antagonistically with DSIF and NELF. In other words, DSIF and NELF bind to RNA polymerase II immediately after transcription initiation and act as a brake to inhibit transcription elongation. P-TEFb (CDK9-Cyclin T) has protein phosphatase activity and phosphorylates the C-terminal domain (CTD) of RNA polymerase II and the C-terminal domain (CTR) of DSIF, thereby inducing the release of NELF and reactivating transcription. It has become clear that P-TEFb-mediated phosphorylation triggers the recruitment of additional protein factors that bind to CTD and CTR and results in the formation of a mature transcription elongation complex.
The field of transcription elongation research has evolved with the identification of numerous transcription elongation factors. Although it has become a well-established field such that university textbooks write about it, it continues to be an active research field with many papers dealing with its functional and structural studies published in top journals.