転写反応はまずRNAポリメラーゼがゲノム上の遺伝子領域にリクルートされる開始段階、RNAポリメラーゼが遺伝子領域を5’方向から3’方向へ向かってmRNA合成を進める伸長段階、RNAポリメラーゼが遺伝子の転写を終えてゲノム上から離れる終結段階の3つの段階に分かれています。これらの反応はゲノムDNAやRNAポリメラーゼに結合する様々な因子の働きによって行われています。1990年代に時計の針を戻すと、当時は転写開始段階の研究が真っ盛りで、真核生物における遺伝子発現のオン・オフ制御は転写開始段階こそが重要と広く信じられていました。
そのような状況下で、当研究室はDRBという転写阻害剤に注目しました。DRBはRNAポリメラーゼII転写の特異的阻害剤で、哺乳動物培養細胞に添加すると、長い転写産物の合成が阻害され、短い転写産物が蓄積することが報告されていました。したがって、DRBは転伸長段階を阻害している可能性が考えられましたが、DRBは意外なことに、精製されたRNAポリメラーゼIIを阻害しませんでした。そこで私達は、生化学的アプローチによりDRBの阻害のメカニズム解明を目指しました。
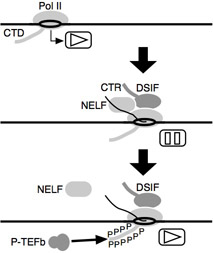 その結果、私達はDSIF(SPT4-SPT5)とNELFという2つの転写伸長因子を新規に同定しました。また、その当時、競っていたDavid Price研が発見したP-TEFbという転写伸長因子が、DSIF、NELFと拮抗的に働くことを突き止めました。すなわち、DSIFとNELFは、転写開始直後のRNAポリメラーゼIIに結合することで、転写伸長を阻害するブレーキ役として働きます。これを解除するのがP-TEFb(CDK9-Cyclin T)です。P-TEFbはタンパク質リン酸化酵素活性をもち、RNAポリメラーゼIIのC末端ドメイン(CTD)や、DSIFのC末端ドメイン(CTR)などをリン酸化することでNELFの脱離を誘導し、転写を再活性化します。このリン酸化をきっかけにして、CTDやCTRに結合するタンパク質因子がさらにリクルートされ、成熟した転写伸長複合体へと変化することが分かってきました。
その結果、私達はDSIF(SPT4-SPT5)とNELFという2つの転写伸長因子を新規に同定しました。また、その当時、競っていたDavid Price研が発見したP-TEFbという転写伸長因子が、DSIF、NELFと拮抗的に働くことを突き止めました。すなわち、DSIFとNELFは、転写開始直後のRNAポリメラーゼIIに結合することで、転写伸長を阻害するブレーキ役として働きます。これを解除するのがP-TEFb(CDK9-Cyclin T)です。P-TEFbはタンパク質リン酸化酵素活性をもち、RNAポリメラーゼIIのC末端ドメイン(CTD)や、DSIFのC末端ドメイン(CTR)などをリン酸化することでNELFの脱離を誘導し、転写を再活性化します。このリン酸化をきっかけにして、CTDやCTRに結合するタンパク質因子がさらにリクルートされ、成熟した転写伸長複合体へと変化することが分かってきました。
これら以外にも多数の転写伸長因子が同定され、転写伸長の研究分野は発展してきました。大学の教科書にも取り上げられるようになり、分野として確立された感はありますが、いまだトップジャーナルに機能解析や構造解析の論文が掲載され、アクティブな研究分野であり続けています。