![]()
トップ > ゲノム発現制御機構
ゲノム発現制御機構
はじめに
当研究室では、ゲノムから機能的RNAが作り出されるまでの過程―すなわちエピジェネティック制御、転写の開始・伸長・終結、RNAプロセシングという一連の過程―の解明を目指しています。私達の体を構成する1つ1つの細胞は、同一のゲノムを持っているにも関わらず多彩な機能を果たしています。これは、ゲノム情報の発現が時空間的に極めて巧妙に制御されているためです。当研究室では、ゲノム情報発現の過程を制御するタンパク質因子群の同定・解析や、ゲノムから多様なトランスクリプトームが生じるメカニズムの解明を目指した研究を行なっています。こうした地道な基礎研究の積み重ねは、一例を挙げればES細胞がES細胞らしい性質を保持するしくみの解明などに役立ちます。
転写伸長制御のメカニズム
転写反応はまずRNAポリメラーゼがゲノム上の遺伝子領域にリクルートされる開始段階、RNAポリメラーゼが遺伝子領域を5’方向から3’方向へ向かってmRNA合成を進める伸長段階、RNAポリメラーゼが遺伝子の転写を終えてゲノム上から離れる終結段階の3つの段階に分かれています。これらの反応はゲノムDNAやRNAポリメラーゼに結合する様々な因子の働きによって行われています。1990年代に時計の針を戻すと、当時は転写開始段階の研究が真っ盛りで、真核生物における遺伝子発現のオン・オフ制御は転写開始段階こそが重要と広く信じられていました。そのような状況下で、当研究室はDRBという転写阻害剤に注目しました。DRBはRNAポリメラーゼII転写の特異的阻害剤で、哺乳動物培養細胞に添加すると、長い転写産物の合成が阻害され、短い転写産物が蓄積することが報告されていました。したがって、DRBは転伸長段階を阻害している可能性が考えられましたが、DRBは意外なことに、精製されたRNAポリメラーゼIIを阻害しませんでした。そこで私達は、生化学的アプローチによりDRBの阻害のメカニズム解明を目指しました。
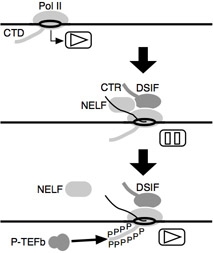
その結果、私達はDSIF(SPT4-SPT5)とNELFという2つの転写伸長因子を新規に同定しました。また、その当時、競っていたDavid Price研が発見したP-TEFbという転写伸長因子が、DSIF、NELFと拮抗的に働くことを突き止めました。すなわち、DSIFとNELFは、転写開始直後のRNAポリメラーゼIIに結合することで、転写伸長を阻害するブレーキ役として働きます。これを解除するのがP-TEFb(CDK9-Cyclin T)です。P-TEFbはタンパク質リン酸化酵素活性をもち、RNAポリメラーゼIIのC末端ドメイン(CTD)や、DSIFのC末端ドメイン(CTR)などをリン酸化することでNELFの脱離を誘導し、転写を再活性化します。このリン酸化をきっかけにして、CTDやCTRに結合するタンパク質因子がさらにリクルートされ、成熟した転写伸長複合体へと変化することが分かってきました。
これら以外にも多数の転写伸長因子が同定され、転写伸長の研究分野は発展してきました。大学の教科書にも取り上げられるようになり、分野として確立された感はありますが、いまだトップジャーナルに機能解析や構造解析の論文が掲載され、アクティブな研究分野であり続けています。
転写伸長制御の生理学的意義
転写伸長制御のメカニズム研究とは別に、その生理学的意義の解明を目指す研究が並行して行われています。ヒト培養細胞をモデル系として用いて行う研究に加えて、様々なモデル生物を用いて共同研究として進める研究もあります。転写伸長制御の生理学的意義としてまず明らかなのは、前初期遺伝子(immediate-early genes)と総称されるような、外部からの刺激に応答して素早く発現する遺伝子群の転写誘導に果たす役割です。つまり、mRNAの完成直前の段階でRNAポリメラーゼIIの転写を止め、クロマチンはアクティブな状態に保っておき、いざ外部から刺激が入ったら速やかに転写を再開できるようにする、というのが転写伸長制御の生理学的意義と考えられ、熱ショック、増殖因子、ホルモン等に応答して発現する遺伝子群を対象とした研究で、実際にこのモデルが確かめられてきました。
また、ゼブラフィッシュやショウジョウバエ個体を用いた共同研究により、転写伸長制御が脳神経系の発生・分化に重要な役割を果たしていることも明らかとなってきました。発生・分化過程では、多数の細胞種特異的遺伝子の発現がダイナミックに変動しているので、前初期遺伝子と似た状況が現出していると考えられます。
さらに、近年発展著しい次世代シーケンサーにより、ゲノム全体の情報が詳細に得られるようになりました。それにより、生化学では絶対にアプローチできない、個々の遺伝子上でのRNAポリメラーゼIIや転写因子の振る舞いを調べることが可能になりました。そして実際、DSIFやNELFなどによる転写伸長制御がゲノム発現全体に重要であることが分かってきました。
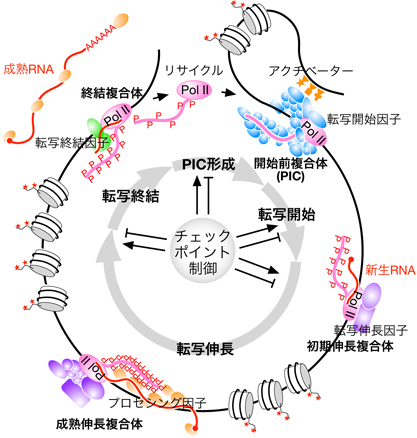
転写終結制御のメカニズム:多様なトランスクリプトームはいかにして生じるのか
当研究室は、転写終結段階の制御によって多様なトランスクリプトームが生じるメカニズムについても積極的に研究しています。RNAポリメラーゼIIが転写する遺伝子には3種類のRNA 3'末端プロセシング経路が存在し、それらが遺伝子によって使い分けられています。また原理上、3'末端プロセシングは転写終結と共役しており、3'末端プロセシングが転写終結を誘導しますが、私達は最近、転写の開始や伸長などに関与すると考えられていた因子(NELF、CBC、Mediator、LECなど)が3種類のプロセシング経路の使い分けにも関与している証拠を得ました。さらに、1つの遺伝子に複数の転写終結点が存在するケースが多々あり、それらの使い分けに関与する因子群についても同定・解析を進めています。同一のゲノムから、細胞種ごとに多様なトランスクリプトームが生じますが、その一端がこうした転写終結段階の制御によって達成されていると考えられます。私達は次世代シーケンサーを用いたゲノム・トランスクリプトーム解析に加えて、最先端の質量分析装置を用いたプロテオーム解析やCRISPR/Cas9を用いた遺伝学的な手法を組み合わせて、これらの課題に取り組んでいます。そこでは、生命情報学のアプローチが大変重要になります。

![]()
- 大岡山キャンパス
- 〒152-8550 東京都目黒区大岡山2-12-1
- すずかけ台キャンパス
- 〒226-8503 神奈川県横浜市緑区長津田町 4259
- 田町キャンパス
- 〒108-0023 東京都港区芝浦3-3-6
![]()
Copyright (C) Tokyo Institute of Technology. All rights reserved.